Emergent Convergence to Euler's Number in Cellular Automata In Progress
Abstract
We present ongoing research into a novel cellular automaton system where purely local interactions between cells drive a global metric toward convergence on Euler's number, e ≈ 2.71828. Our experiments demonstrate that the introduction of spatial barriers (walls) can significantly influence convergence dynamics, with partial barriers often improving convergence rates compared to unrestricted systems. These findings suggest potential pathways toward understanding how intelligent-like behavior might emerge from simple local rules operating under physical constraints.
Introduction and Motivation
The emergence of complex, seemingly purposeful behavior from simple rules is a central question in artificial life, complexity science, and the foundations of intelligence. Cellular automata have long served as laboratories for studying emergence—from Conway's Game of Life to Wolfram's elementary automata.
Our research asks a more targeted question: Can local cellular interactions, without any global coordination, cause a distributed system to converge on a specific mathematical constant?
We focus on Euler's number e, chosen for its fundamental role in mathematics and its representation as an infinite series:
e = 1 + 1 + 1/2 + 1/6 + 1/24 + ... ≈ 2.71828
The long-term vision is to understand whether such convergent behavior could serve as a primitive form of "goal-directed" computation—a stepping stone toward systems that exhibit intelligent behavior through emergent dynamics rather than explicit programming.
The Cellular Space Model
Our system consists of a 2D grid of cells, each holding a positive integer value. In our standard configuration we use a 10 × 10 grid (100 cells) with initial values randomly distributed in [1, 100], and an 8-connected neighborhood (Chebyshev distance, like a chess king).
At any moment, we compute the inverse sum metric:
S = 1 + Σ(1/vi) for all cells
This metric is inspired by the series representation of e. As cell values grow large, their reciprocals approach zero, and the sum stabilizes. The question is: toward what value?
Each simulation proceeds in rounds. During a round, every active cell performs one operation: (1) inspect neighbors within a configurable distance, (2) identify the neighbor with the largest value, (3) multiply that neighbor's value by the acting cell's value, and (4) swap positions with the target cell.
This creates a dynamic where values grow exponentially through multiplication, spatial positions evolve as cells "chase" high-value targets, and the inverse sum metric changes with each operation.
Baseline Convergence Behavior
We track the convergence metric after every individual cell operation, not just at round boundaries. This reveals the fine-grained dynamics of approach toward e.
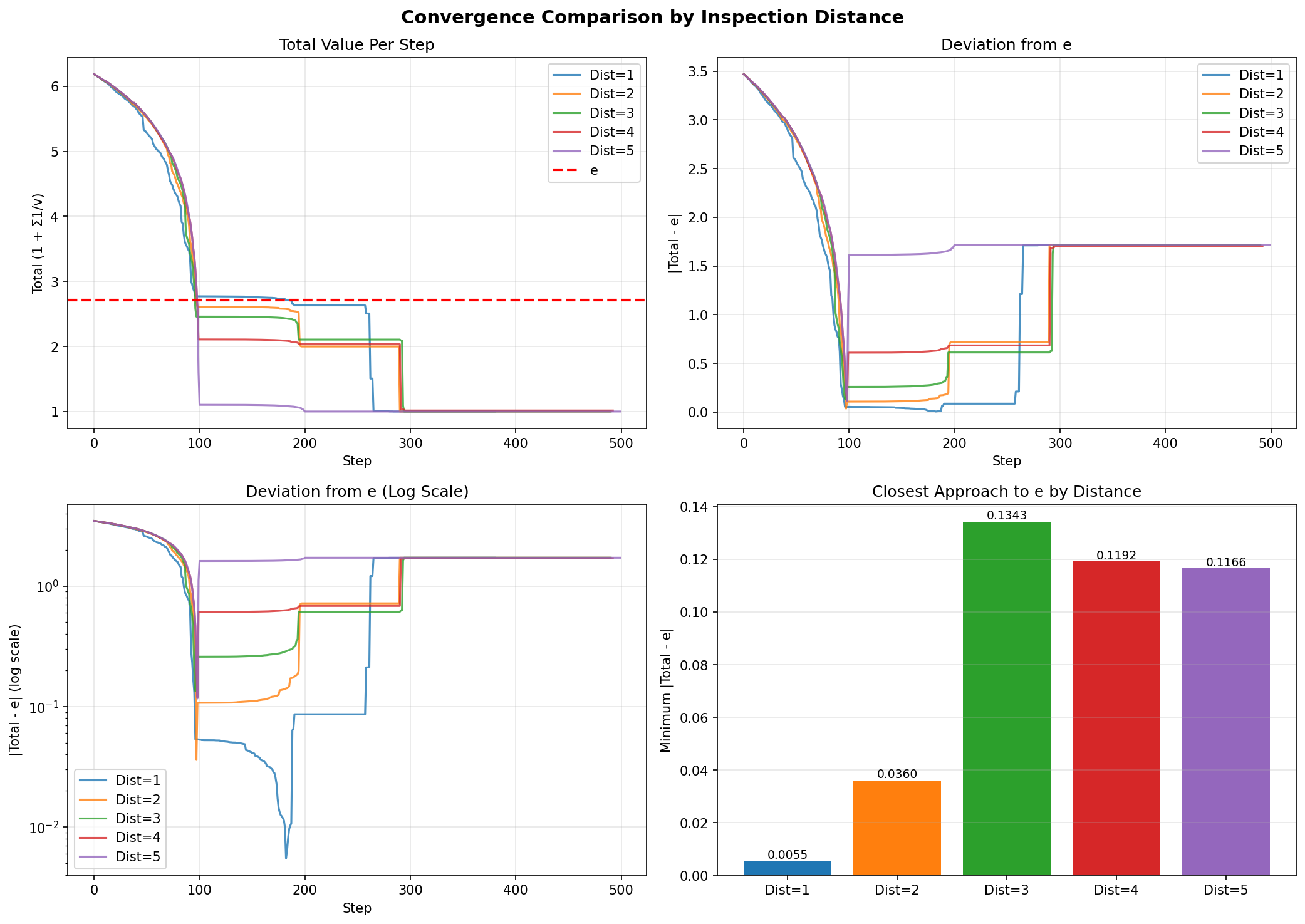
Figure 1: Convergence behavior across different inspection distances. Distance 1 (immediate neighbors only) achieves the closest approach to e.
| Distance | Closest Approach | Deviation from e |
|---|---|---|
| 1 | 2.7128 | 0.0055 |
| 2 | 2.6823 | 0.0360 |
| 3 | 2.5840 | 0.1343 |
| 4 | 2.5990 | 0.1192 |
| 5 | 2.6016 | 0.1166 |
Finding: Smaller inspection distances yield better convergence. This is counterintuitive—one might expect that seeing more neighbors would help. Instead, the locality of interactions appears crucial. All configurations eventually stagnate around round 4-5, when cell values have grown so large that further operations produce negligible changes to the inverse sum.
Wall Experiments: Spatial Constraints and Convergence
If local interactions drive convergence, how do spatial barriers affect the dynamics? We hypothesized that walls—which block both line-of-sight and cell movement—would create isolated subsystems with potentially different convergence properties.
Walls are placed on edges between cells, not within cells. They affect the system in two ways: (1) line-of-sight blocking via ray-casting, and (2) movement blocking that prevents cell swapping across walls.
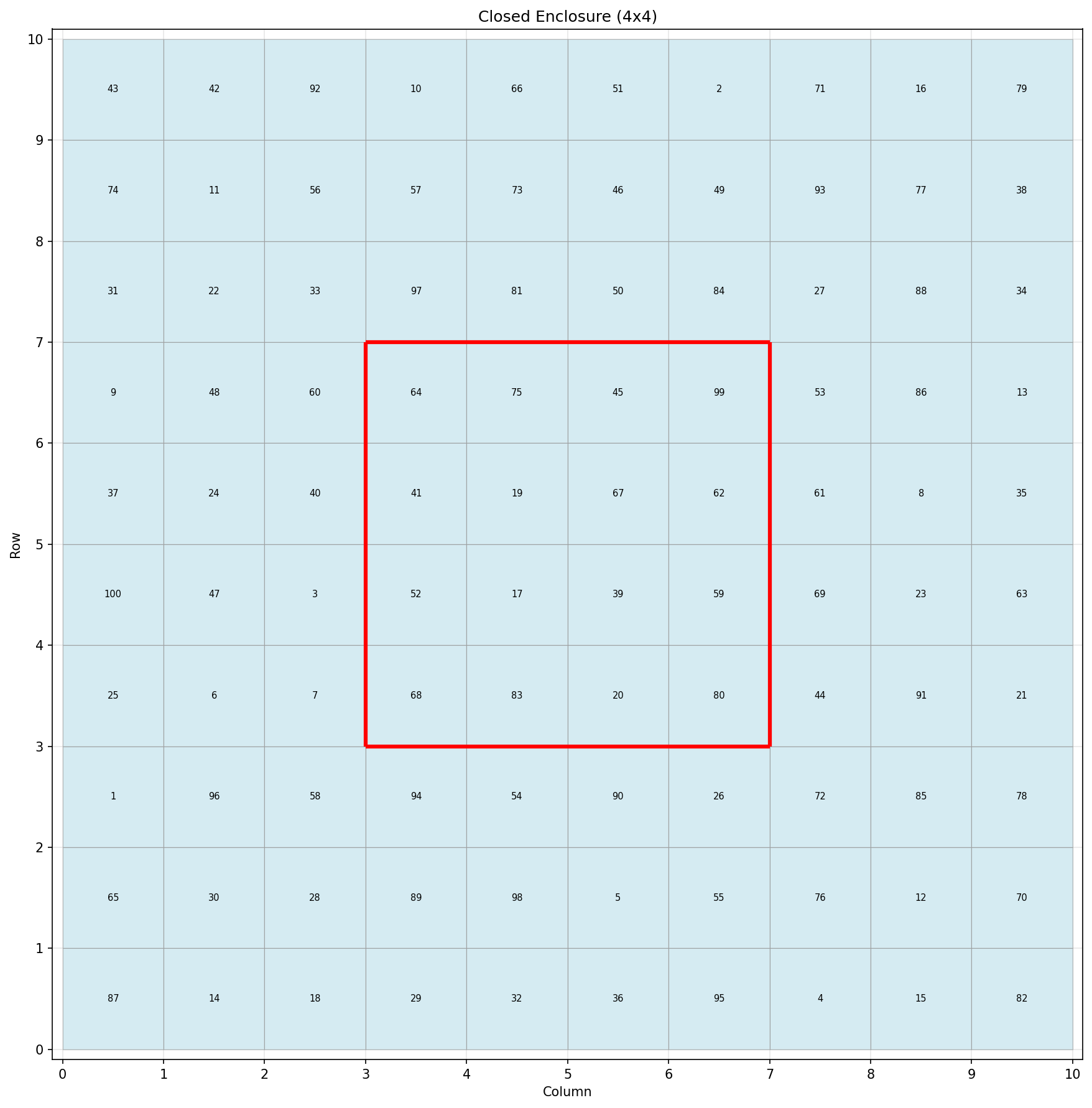
Figure 2: Example wall configuration—a 4×4 enclosed region that is completely isolated from the surrounding grid.
We tested seven wall configurations: No Walls (baseline), Horizontal Wall (across grid center), Vertical Wall (down grid center), L-Shape (in one quadrant), T-Shape (from top), Enclosure (completely isolated 4×4 box), and Random (8 randomly placed segments).
Statistical Results
We ran 50 iterations per configuration with different random seeds to obtain statistically meaningful results.
| Configuration | Mean Min Dev | Std Dev | Rank |
|---|---|---|---|
| H-Wall | 0.1008 | 0.109 | 1 (best) |
| V-Wall | 0.1021 | 0.115 | 2 |
| T-Shape | 0.1050 | 0.119 | 3 |
| No Walls | 0.1113 | 0.125 | 4 |
| Random | 0.1143 | 0.135 | 5 |
| L-Shape | 0.1155 | 0.132 | 6 |
| Enclosure | 0.1464 | 0.148 | 7 (worst) |
Key Findings
Finding 1: Partial barriers improve convergence. Both horizontal and vertical walls through the center of the grid yielded better average convergence than the unrestricted baseline. This suggests that some degree of spatial constraint creates beneficial dynamics—perhaps by allowing subsystems to evolve at different rates before interacting.
Finding 2: Complete isolation hurts convergence. The enclosed region performed worst. When cells are completely cut off from the larger system, they lack the diversity of interactions needed to drive the inverse sum toward e. The isolated 4×4 region and the surrounding 84 cells each converge to their own local equilibria, which don't combine to approximate e.
Finding 3: The topology matters. The specific geometry of barriers affects outcomes. T-shaped walls (which create three semi-isolated regions) outperformed L-shaped walls (which create two). This hints at an optimal "mixing" regime between full connectivity and full isolation.
Flow Visualization
To better understand the spatial dynamics, we developed flow visualizations tracking how values move through the grid over time.
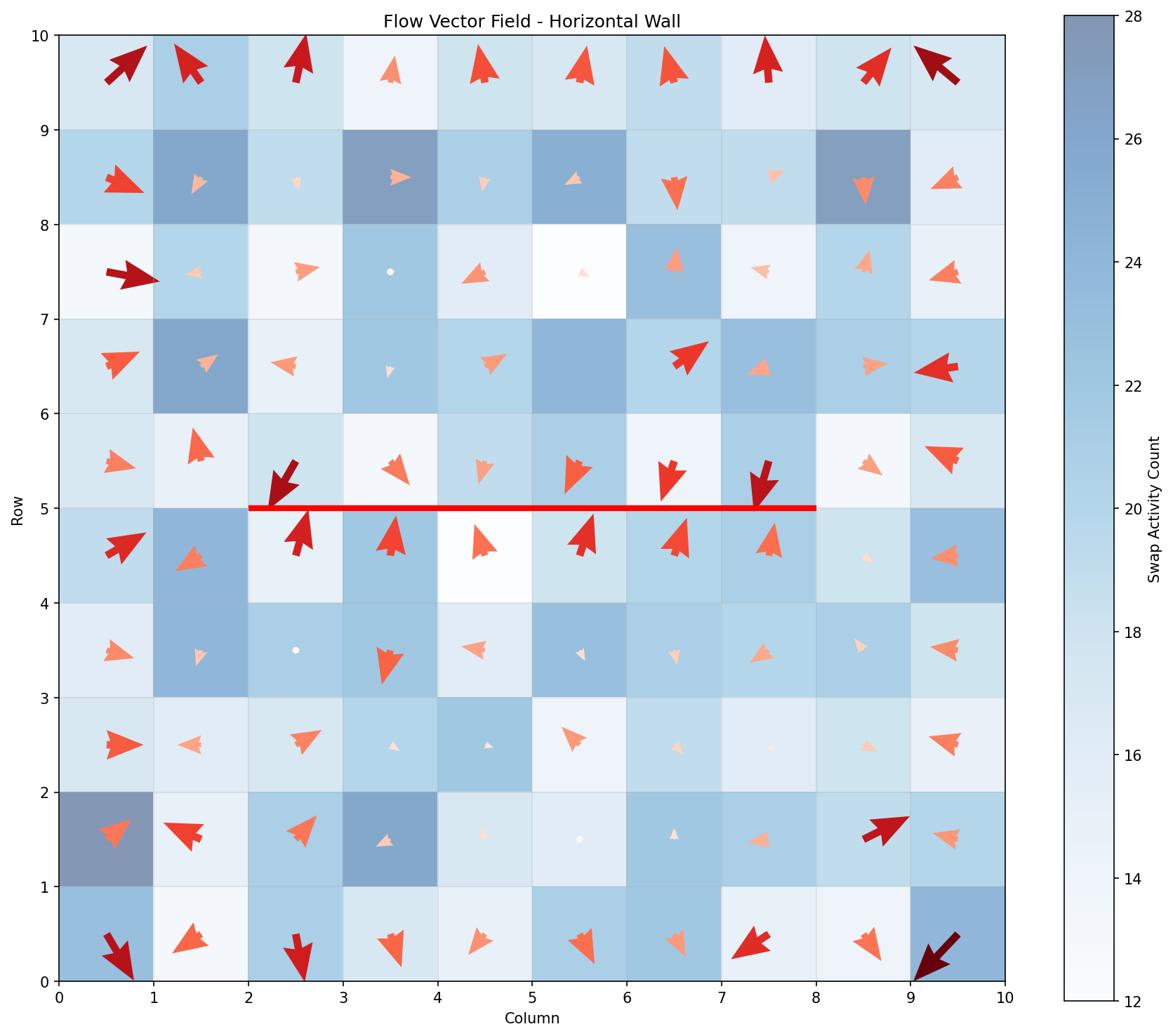
Figure 3: Flow vector field for horizontal wall configuration. Arrows show average movement direction; heatmap shows swap activity intensity.
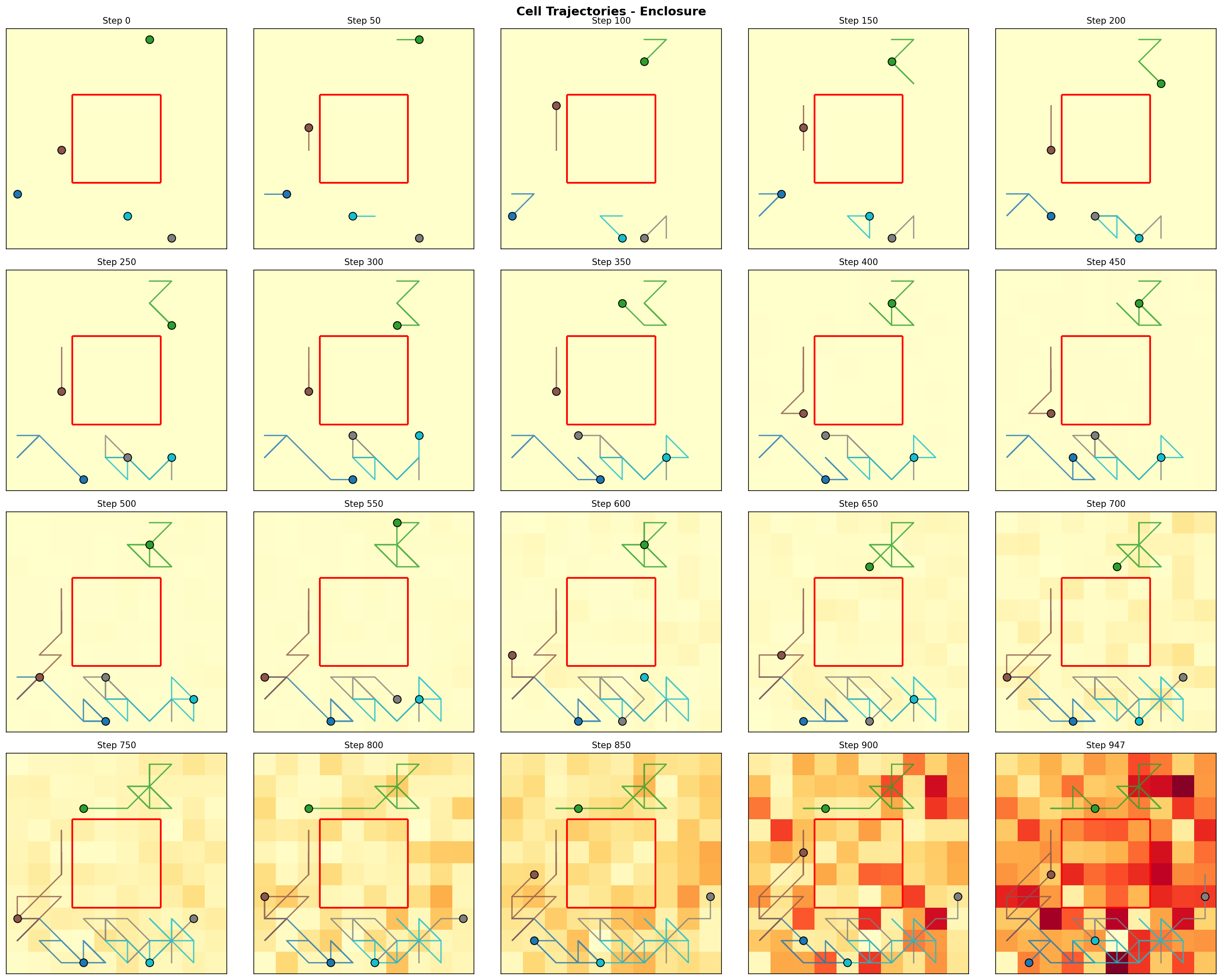
Figure 4: Trajectory snapshots for the enclosure configuration. Cells inside the enclosed region remain trapped, while outside cells move freely.
Discussion
The convergence toward e emerges from the multiplicative growth dynamics. When cell A multiplies cell B's value, the reciprocal of the product is the product of reciprocals. This creates a cascading effect where the inverse sum shrinks in a structured way. The specific dynamics—which cells interact, in what order—determine the final convergence point.
Our long-term research program asks whether convergent cellular dynamics could exhibit properties associated with intelligence: goal-directedness (the system moves toward a specific target without explicit programming), adaptation (different initial conditions still lead toward similar endpoints), and robustness (partial barriers don't destroy convergence—they modulate it).
These are preliminary observations, not claims of intelligence. But they suggest a research direction: studying how spatial constraints and local rules can produce globally coherent, seemingly purposeful behavior.
Limitations
A fundamental limitation of the current system is its inability to stabilize at e. While the inverse sum metric approaches e during the simulation, it does not rest there—it continues past e and ultimately converges toward 1.
This behavior is an inevitable consequence of the multiplication rule. Each operation increases at least one cell's value, which decreases its reciprocal contribution to the sum. Since there is no mechanism to decrease cell values or halt multiplication, the inverse sum can only shrink monotonically. The system passes through e as a transient state rather than reaching it as an equilibrium.
Mathematically, as all cell values approach infinity, the metric approaches its lower bound:
lim S = 1 + Σ(1/∞) = 1 + 0 = 1
This represents a critical gap between the current implementation and the goal of emergent convergence to e. A system that genuinely "targets" e would require rules that create a stable equilibrium at that value—where the dynamics naturally balance rather than overshooting. More work is needed to determine a suitable rule set that allows the system to naturally express the converging series and rest at e rather than merely passing through it.
Potential directions include: introducing division or decay operations that counterbalance multiplication, implementing energy-like conservation laws that constrain total growth, or designing feedback mechanisms where cells respond to the global metric. These modifications would fundamentally change the system's character and require careful analysis to preserve the emergent, locally-driven nature of the dynamics.
Open Questions
Several questions remain open for future investigation: Why e specifically? Is there something special about our rules that targets e, or would different rules target different constants? What wall configuration maximizes convergence? Is there a principled way to design barriers? How does convergence behavior change with grid size or with 3D grids? Can we characterize the "information" that crosses (or is blocked by) walls?
Current Status and Next Steps
This work is ongoing. We have a working simulation framework with wall support, a statistical analysis pipeline (50+ iterations per configuration), and flow visualization tools (vector fields, trajectory animations). We have preliminary evidence that spatial constraints affect convergence.
Next steps include: developing mathematical models explaining why partial barriers help, searching for wall configurations that maximize convergence to e, testing whether modified rules can target other constants (π, φ, etc.), defining and measuring "intelligent-like" properties more rigorously, and testing on larger grids and higher dimensions.
Conclusion
We have demonstrated that a simple cellular automaton with local multiplicative interactions naturally converges toward Euler's number e. More surprisingly, introducing spatial barriers—walls that block sight and movement—can improve convergence compared to unrestricted systems, provided the barriers are partial rather than complete.
These findings open questions about the relationship between spatial structure, local dynamics, and emergent global behavior. While we are far from claims about intelligence or consciousness, we believe this line of research offers a novel approach to understanding how goal-directed behavior might arise from simple, distributed rules operating within physical constraints.